|

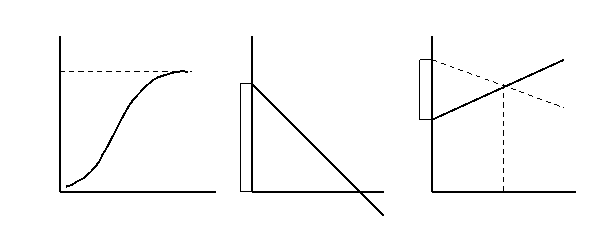 
а r б b, d в
d
K
rmax
b
r max
N0 N N
0 Время, t 0 K 0 K
Рис. 3. Логистическая модель роста популяции: а – кривая роста численности (N); б – зависимость удельной скорости роста (r) от численности (N); в – зависимость рождаемости (b) и смертности (d) от численности; К – предельная численность.
В природе так и происходит: экспоненциальный рост наблюдается какое-то достаточно короткое время, после чего ограничивающие факторы его стабилизируют, и дальнейшее развитие популяции идет по логистической модели, что и описывается S – образной, или логистической кривой роста популяции.
В основе логистической модели (рис. 3) лежит простое допущение, что скорость роста популяции (ra) линейно снижается по мере роста численности вплоть до нуля при некой численности К. Итак, при начальной численности Na (близкой к нулю) скорость роста имеет максимальное значение r max , а при N = K, r a = 0. В результате решения уравнения логистической кривой получается зависимость:
Nt = K / (1 + e α – r max ∙ t ),
где Nt – численность популяции в момент времени t;
α – постоянная интегрирования.
Величины К называют еще емкостью среды в отношении особей данной популяции. Здесь речь идет о биологической емкости среды – степени способности природного или природно-антропогенного окружения обеспечить нормальную жизнедеятельность (дыхание, питание, размножение, отдых и т.д.) определенному числу организмов и их сообществ без заметного нарушения самого окружения.
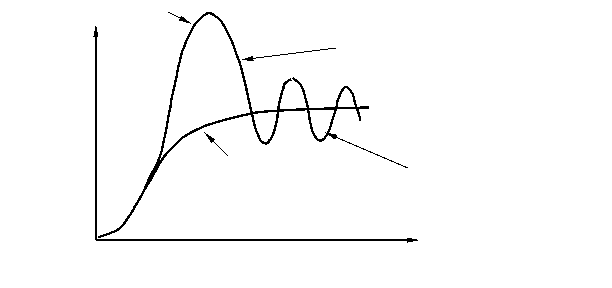
Однако плато на S – образной кривой далеко не всегда бывает гладким, потому что колебания численности происходят постоянно, что отражается в виде колебаний кривой вокруг асимптоты К (рис. 4), эти колебания называются популяционными волнами или флюктуациями численности. Данный термин был введён русским биологом С. С. Четвериковым в 1915 г. Подобные колебания численности могут быть сезонными или несезонными, повторяющимися через различные промежутки времени; обычно они тем длиннее, чем продолжительнее цикл развития организмов. Часто волны жизни сопровождаются колебаниями ареала популяций. Четвериков указал на эволюционное значение волн жизни в качестве фактора, могущего изменять направление и интенсивность отбора, а также концентрации генов, содержащихся в популяциях. Популяционные волны (один из 4 элементарных эволюционных факторов — мутационный процесс, популяционные волны, изоляция и естественный отбор). Основное значение популяционных волн сводится к случайным изменениям концентраций (особенно невысоких) различных мутаций и генотипов, содержащихся в популяциях, а также к ослаблению давления отбора при увеличении и его усилению при уменьшении численности особей в популяции. Под термином «популяционные волны» иногда (советский геолог Б. Л. Личков и некоторые другие ученые) также подразумевают этапы развития растительного и животного мира, примерно соответствующие смене геологических циклов.
Повышение популяцией
потенциальной емкости
«Крах» популяции
или вымирание
Число
особей
Потенциальная Популяционные
емкость волны
Время
Рис. 4. Преобразование J –образной кривой роста численности популяции в S – образную кривую при ограничивающем воздействии лимитирующих факторов.
Главенствующее значение для естественного возобновления ценопопуляции имеет запас диаспор в почве, наличие подроста и занос зачатков. В ненарушенной лесной среде практически исключена смена лесного типа растительности на луговой, степной или болотный. Если же ширина окна превышает две и более высоты сомкнутого окружающего полога, то возникает нелесная среда, где преимущество заселения получают вначале травяной и моховой типы растительности, и только потом заселяются пионерные береза, осина, иногда сосна, при этом частично или навсегда исчезают многие лесные виды растений, животных и, вероятно, микроорганизмов.
Можно наблюдать следующие закономерности возникновения возобновления сообществ. Почвенный запас семян хорошо защищен от поражения микроскопическими грибковыми организмами, неспорообразующими бактериями, спорообразующими бактериями, актиномицетами, но недостаточно – от представителей лесной фауны, особенно видов, питающихся в основном семенами. Случайным является попадание семян в оптимальные условия прорастания, совпадающие с генетической наследственностью. При прорастании семени проросток сразу вступает в тесные взаимоотношения с микробиоценозом, эдафотопом и климатопом, т.е. любое растение образует с биоценозом консорцию. В этот период наблюдается массовая гибель растений из-за несовпадения консорциативных и неблагоприятных экотопических связей. Объем пространства, занимаемый проростками и ювенильными древесными растениями, невелик, и конкуренции за свет и почву между ними нет. Ресурсы среды у них могут отнять мхи, травы и взрослые древесные особи. Это предопределяет всегда недостаточное число возобновления на начальном этапе. Даже количество возобновления, равное 2,5 млн. шт./га, означает 40 см2 (5 x 8 см), приходящихся на одно растение. Следовательно, энергию роста (бонитет) на начальном этапе полностью определяют почвенное богатство и сопряженность с микробоценозом. В это время закладывается жизненная стратегия роста и развития (так называемые классы Крафта – деление деревьев в лесу на пять классов по росту и развитию: прегосподствующие, господствующие, умеренно развитые, заглушенные и подпологовые) каждой особи. По мере роста особь увеличивает занимаемое ею пространство до начала смыкания кронами с другими особями, после чего начинается закономерная межвидовая и видовая ценопопуляционная конкуренция между особями в биогруппе вначале за свет надземного пространства, позже присоединяется конкуренция корневых систем особей за почвенные ресурсы. По отношению к каждой особи этот процесс получил название «давление среды», приводящее к изреживанию в биогруппах и в насаждении в целом. Выжившие особи увеличивают консорциативные связи, вступая в тесные взаимоотношения со всем биоценозом, формируя собственную свиту растений, животных, грибов, микроорганизмов и изменяя внутреннюю среду биогеоценоза в выгодном для себя отношении. В метрическом исчислении это выражается двумя показателями – высотой и занимаемым воздушным и почвенным объемами, т. е. числом особей на 1 га. Чем беднее почва, тем больше необходимый занимаемый особью объем почвы, в связи, с чем площадь крон отстает от площади корневой системы, в результате исключается стопроцентная сомкнутость крон, а значит не полностью создается лесная среда. Особенно это видно в переходных типах растительности. При любом возрасте конкурентные отношения выражаются высотой и числом стволов, что позволяет сравнивать с высокой степенью корреляции эти показатели при разных бонитетах и возрастах.
|