|

Ценотическая регуляция филогенеза. Эта модель, как отмечалось, является дальнейшим развитием идей В. В. Жерихина. В его работах главное внимание обращено на регулирующее воздействие структуры адаптивных зон и экологических ниш сформированного сообщества на процессы филогенеза, входящих в его состав — ценофильных таксонов (Жерихин, Расницын, 1980; Жерихин, 1987).
Такую ценотически регулируемую эволюцию Жерихин вслед за В. А. Красиловым (1969, 1989) называет когерентной. Природу срединномелового биоценотического кризиса, обнаруженного и проанализированного Жерихиным (1978—1987), он объясняет снятием ценотических ограничений филогенеза вследствие разрушения структуры адаптивных зон и экологических ниш биоценоза, и переходом эволюции таксонов в некогерентную фазу. Снятие ограничений обеспечивает быструю в геологическом масштабе времени перестройку сообщества растений, насекомых (Жерихин, 1978, 1980) и ряда других групп животных суши (Жерихин, 1979). Аналогичное событие в ранней—средней юре, но избирательно коснувшееся только сообщества наземных тетрапод, которому посвящена первая часть статьи, мы склонны объяснить так же. Именно события подобного рода, хотя и косвенно, убеждают в существовании и эффективности ценотической регуляции филогенеза в сформированном сообществе.
Однако, если причина кризиса внутренняя, то, указав ближайшую — непосредственную его причину, остается открытым вопрос о процедуре ее исторической подготовки. Вариант ответа на этот вопрос, в частности, предлагает рассматриваемая ниже модель.
Наша задача обратить внимание на противоположный процесс: регуляцию эволюции сообщества процессами филогенеза таксонов. Такое дополнение представлений Жерихина позволяет предложить модель саморазвития сообщества (филоценогенеза) и таксономического разнообразия биоты (филогенеза) по принципу обратной связи. При этом противоречивые отношения в системе сообщество—биота оказываются в конечном счете главной движущей силой их сопряженного развития.
Материал. В основу оригинальной части нашего построения легли распространенные в палеонтологической летописи феномены динамики таксономического разнообразия разных, но экологически сопряженных групп организмов, составляющих одно широко понимаемое сообщество в процессе его исторического развития. Типичная последовательность этих феноменов схематически представлена в левой части рис. 4. История сообщества начинается явным преобладанием относительно малоспециализированных форм, разнообразие которых резко сокращается по мере прогрессивной специализации его членов. Последнее сопровождается дивергенцией и ростом таксономического разнообразия. Затем разнообразие специализированных форм начинает сокращаться без видимых причин, т. е. в отсутствие явно превосходящего по экологической валентности конкурента, и на этом фоне начинает вновь расти разнообразие малоспециализированных форм. Предположение о конкурентном вытеснении первых вторыми является очевидным экологическим нонсенсом. И наконец, в довершение всего возникает новая формация специализированных таксонов, на фоне роста разнообразия которых продолжает сокращаться разнообразие специалистов прежней формации и вновь уменьшается разнообразие малоспециализированных форм. При этом, по крайней мере в начале процесса, первые должны уступать по уровню специализации вторым и потому не могли их вытеснять конкурентно. Это дополнительно подтверждается тем, что сокращение разнообразия специалистов первой формации в подобных случаях начинается задолго до того, как специалисты второй формации становятся многочисленны и разнообразны.
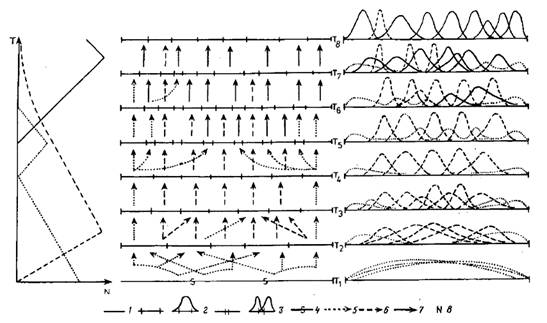
Рис. 4. Модель сопряженной эволюции сообщества, его таксономического и экологического разнообразия: 1 — пространство ресурсов сообщества, 2 — адаптивная зона, 3 — интерзональная экологическая лакуна, 4 — нечетко выраженная граница адаптивных зон, 5 — слабо специализированный таксой, 6—высоко специализированный таксон первой формации, 7—высоко специализированный таксон второй формации, 8—мера таксономического разнообразия
Вся картина в целом начинается экологически очевидным феноменом конкурентного вытеснения малоспециализированных форм более специализированными, но далее приобретает характер процесса, не имеющего тривиальной экологической интерпретации. Приведем два подобных примера из нашей практики.
Перейти на страницу: 1 2 3 4 5 6 |