|

1 -й пример. На рис. 5, а показана динамика таксономического разнообразия плотоядных зверей Северной Америки. Часть результатов ее анализа была опубликована раньше (Раутиан, 1980). По мере роста разнообразия специализированных хищников палеогена (в крупном размерном классе: Arctocyonia и Crcodonta; в среднем: Miacidae; в мелком — древние группы насекомоядных с относительно низким уровнем специализации: Pantolestidae, Adapisoricidae, Solenodontia, Leptictida) резко сокращается разнообразие самых низко специализированных плотоядных — Didelphoidea, доминировавших в мелу. Их разнообразие вновь подскакивает в олигоцене на фоне сокращения разнообразия перечисленных специалистов. Вслед за этим доминирующими специализированными хищниками становятся Feloidea и Canoidea, занимающие постепенно все размерные классы хищников, а специализированными насекомоядными, так или иначе ушедшими от прямой конкуренции с хищниками (в отличие от более древних вытесненных групп), становятся Erinaceidae, Talpidae и Soricidae. Они составляют вторую формацию специалистов. Падение разнообразия специалистов первой формации начинается задолго до того, когда специалисты второй формации становятся многочисленны и разнообразны. Именно это позволяет легко различить специалистов разных формаций. Таким образом, смену специалистов разных формаций не удается объяснить посредством конкурентного вытеснения. Далее, на фоне роста разнообразия специалистов второй формации, достигшего максимума в миоцене, вымирают сумчатые (Didelphidae). Однако уже в плейстоцене на фоне снижения разнообразия специалистов опоссум (Didelphis) проникает из Южной Америки в Северную, где этот ценофоб в наше время в условиях хозяйственной деятельности человека расширяет географическую область обитания и экологическую сферу своей активности, являясь одновременно вредителем и не очень ценным пушным зверьком.
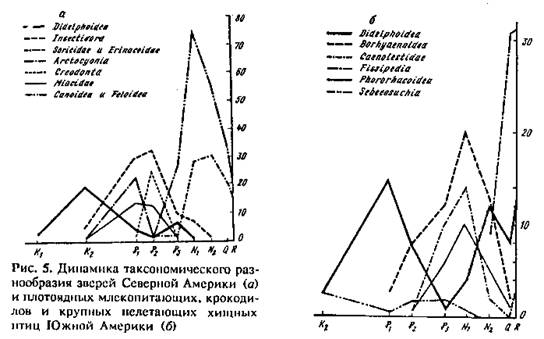
2-й пример, кратко изложенный нами раньше (Каландадзе, Раутиан, 1979), касается плотоядных Южной Америки (рис. 5, б). В мелу и палеогене здесь тоже доминировали Didelphoidea. Позже их сменили в качестве экологических доминантов специализированные хищные (Borhyaenoidea) и насекомоядные (Caenolestidae) сумчатые. Ввиду продолжительного для этого материка дефицита крупных хищных зверей (Каландадзе, Раутиан, 1989) к ним добавились крупные наземные хищные крокодилы (Sebecidae) и многочисленные крупные бегающие хищные птицы (Phororhacoidea). Впечатляющий вторичный рост разнообразия Didelphoidea объясняется прежде всего экологическим кризисом, связанным, по терминологии В. В. Жери-хина, с коллизией биот: внедрением в длительно (с мела, см. рис. 1) изолированную страну (Каландадзе, Раутиан, 1982—1989; Раутиан, 1982) фауны из Северной Америки, обладающей значительно большей экологической валентностью. Это свидетельствует о росте разнообразия малоспециализированных форм в эпоху кризиса. Однако рост разнообразия Didelphoidea, падение разнообразия специализированных хищников и насекомоядов, а также вымирание наземных крокодилов началось задолго до внедрения в Южную Америку североамериканских зверей.
Постановка задачи. Таким образом, предлагаемая модель должна разрешить по крайней мере три экологических парадокса: 1) падение разнообразия специализированных таксонов в отсутствие конкурентов, превосходящих их по экологической валентности; 2) вторичный рост разнообразия малоспециализированных таксонов на фоне высокого разнообразия специалистов; 3) смену специализированных таксонов первой формации таковыми второй формации при условии, что по крайней мере в начале процесса вторые должны уступать по уровню специализации первым и потому не могли их вытеснять конкурентно.
К этому добавляется возникшая еще при чтении первой части статьи проблема исторической подготовки крупного экологического кризиса, которую в первом приближении удается обсудить в связи с решением экологических парадоксов.
Типичная последовательность стадии эволюции сообщества схематически показана на рис. 4. Левая часть его уже рассматривалась выше. В средней части внимание обращено на разделение пространства ресурсов между адаптивными зонами разных таксонов, на характер границ этих зон и преемственную связь (стрелки) обладателей зон на предшествующей и последующей стадиях эволюции сообщества. В правой части метод реализованной экологической ниши (Одум, 1975) применен для характеристики адаптивных зон и отражает величину конкурентного давления (фактического или потенциального), осуществляемого таксонами — обладателями этих зон в разных областях пространства ресурсов.
Перейти на страницу: 1 2 3 4 5 6 7 |